


Inoculación con microorganismos con efecto promotor de
crecimiento (pgpm) en trigo. Conocimientos actuales y experiencias realizadas en
la región pampeana argentina
Ing. Agr. (MSc)Gustavo N. Ferraris
Área de Desarrollo Rural INTA EEA Pergamino. Av Frondizi km 4,5 (2700) Pergamino
El uso de inoculantes biológicos incorporados como tratamientos de semilla
con microorganimos promotores del crecimiento vegetal tales como Pseudomonas sp.
Azospirillum sp., Micorrizas u otros muestra un creciente interés no sólo en
estudios de investigación sino también en evaluaciones extensivas y usos
comerciales en diferentes cultivos. Efectos como una más rápida implantación,
mayor crecimiento de raíces, tolerancia mejorada a patógenos, fijación biológica
no simbiótica de nitrógeno y solubilización de nutrientes son habitualmente
reportados en estas experiencias (Caballero Mellado et al. 1992). Dado el
crecimiento en los costos de producción, las mejoras derivadas de una mayor
eficiencia de uso de los nutrientes y otros recursos a partir de los aportes de
estos tratamientos biológicos serían de relevancia (Ferraris et al., 2008).
El objetivo de este artículo es realizar una revisión de la información generada
sobre los efectos de los principales microorganismos considerados como PGPM
(microorganismos promotores del crecimiento vegetal) en cultivos de gramíneas, y
evaluar su interacción con las prácticas agronómicas utilizadas bajo un criterio
de agricultura sustentable en la Región Pampeana Argentina.
Azospirillum spp
El Azospirillum es por lejos el PGPM más estudiado en gramíneas, reportándose
las primeras experiencias hace más de treinta años (Döbereiner et al. 1977). En
la actualidad, se han identificado 12 especies de Azospirillum spp, aunque en la
producción de inoculantes comerciales se han utilizado A. brasilense y A
lipoferum, siendo la primera la más común a nivel mundial, y la preferida en
Argentina (Puente y Perticari, 2006).
Algunos antecedentes muestran efectos en la fijación libre del nitrógeno
atmosférico, la producción y liberación de hormonas promotoras del crecimiento
radical (ej. auxinas, giberelinas, citoquininas), y de enzimas tales como las
pectinolíticas, distorsionando la funcionalidad de células de las raíces y el
aumento en la producción de exudados (Okon y Labandera-González, 1994).
Asimismo, de manera indirecta, la inoculación con Azospirillum spp podría
promover la proliferación y establecimiento en la rizósfera de otros
microorganismos favorables para el cultivo (Russo et al. 2005).
El Área de Desarrollo Rural de INTA Pergamino realizó entre 2003 y 2007 18
ensayos bajo condiciones habituales de producción, utilizando inoculantes de
diferentes marcas comerciales. Como resultado, se obtuvieron diferencias
significativas en ocho experimentos P<0,10. La respuesta media alcanzó a 283 kg
ha-1, representando un 6,4 % de incremento. Si bien no se pudo establecer una
función que relacione la respuesta a la inoculación con Azospirillum y las
variables de suelo o cultivo evaluadas, un análisis a través de un gráfico
biplot nos permite definir marcadas tendencias (Figura 1). El eje horizontal (1)
discrimina variables de suelo, clima y los rendimientos. El eje vertical (2) por
su parte, discrimina la respuesta a la práctica (abajo) de las precipitaciones
(arriba). Por otra parte, los puntos azules representan cada uno de los
dieciocho experimentos, y los vectores a las variables analizadas. Los vectores
orientados en un mismo sentido indican parámentos asociados de manera positiva,
siendo la correlación mas fuerte cuanto más cerrado sea el ángulo entre ambos
vectores. En cambio, los orientados en sentido contrario indican una asociación
fuerte pero inversa. Cuando dos vectores forman un ángulo recto, las variables
que representan no están asociadas. De esta manera, se observa que la respuesta
absoluta expresada en kg ha-1 (Rta kg/ha), se relacionó en forma estrecha e
inversa con las lluvias totales y las ocurridas entre octubre y noviembre, más
que con ninguna variable. El incremento de respuesta asociado a menores
precipitaciones se basa en la ventaja competitiva para la adquisición de agua y
nutrientes que tendrían los tratamientos inoculados, otorgada por un mayor
crecimiento aéreo y radicular inicial temprano. Por otra parte, en este grupo de
ensayos no se observó relación alguna entre respuesta y el contenido de materia
orgánica (MO), N (kg h/ha) o P (P) en suelo (Figura 1) o los niveles de
nitrógeno (N) y fósforo (P) agregados por fertilización, evidenciando un
comportamiento aditivo ante diferentes condiciones de cultivo. En la mayoría de
estos experimentos, la respuesta fue similar bajo diferentes niveles
tecnológicos, dosis de fertilizante o genotipo (Díaz Zorita et al., 2004; Díaz
Zorita y Fernández Caniggia, 2008, Ferraris et al., 2008) (Figura 2).
Tabla 1: Descripción de un grupo de ensayos de respuesta a Azospirillum spp en Trigo, en el centro-norte de Buenos Aires y Sur de Santa Fe entre los años 2003 y 2007. En la primer columna se expresa la localidad de ensayo: Bo (Bolivar), Ln (Lincoln), SA (San Antonio de Areco), Pe (Pergamino), Arr (Arrecifes) y Ww (Wheelwright).
|
Sitio |
MO |
P_suelo |
N_suelo |
P_dosis |
N_dosis |
pp Jun-Set |
pp Oct_Nov |
pp_totales |
Rsitio |
Rta (Kg/ha) |
Rta (%) |
P= |
|
Bo 03 |
3,1 |
14 |
85,8 |
12 |
69 |
|
|
|
2845,5 |
193 |
6,8 |
P=0,0003 |
|
Arr 05 |
3,3 |
85 |
70 |
16 |
63 |
175 |
173 |
348 |
3207,5 |
83 |
2,6 |
P>0,10 |
|
Bo1 04 |
1,7 |
11 |
55 |
0 |
0 |
140 |
240 |
380 |
4152,5 |
133 |
3,2 |
P>0,10 |
|
Bo2 04 |
1,7 |
11 |
55 |
10,35 |
8,1 |
140 |
240 |
380 |
4852 |
200 |
4,1 |
P>0,10 |
|
Ln 04 |
3,2 |
10 |
24 |
23 |
95 |
150 |
153 |
303 |
4676,4 |
142,3 |
3,0 |
P>0,10 |
|
Pe 04 |
2,68 |
14 |
18 |
11 |
80 |
90,1 |
153,3 |
243,4 |
3611,1 |
3,6 |
0,1 |
0,98 |
|
SA1 04 |
3,02 |
14 |
60 |
12 |
72 |
115 |
182 |
297 |
3833,85 |
140,1 |
3,7 |
P>0,10 |
|
SA2 04 |
3,02 |
14 |
60 |
12 |
72 |
115 |
182 |
297 |
4176,25 |
196,7 |
4,7 |
P>0,10 |
|
Pe1 05 |
2,94 |
12 |
32 |
0 |
0 |
138,3 |
173,9 |
312,2 |
4459,75 |
990,5 |
22,2 |
0,08 |
|
Pe2 05 |
2,94 |
12 |
32 |
20 |
58 |
138,3 |
173,9 |
312,2 |
5255,25 |
-165,5 |
-3,1 |
0,08 |
|
Pe3 05 |
2,94 |
12 |
32 |
20 |
118 |
138,3 |
173,9 |
312,2 |
5185,25 |
302,5 |
5,8 |
0,08 |
|
Pe4 05 |
3,02 |
6 |
32 |
0 |
0 |
138,3 |
173,9 |
312,2 |
4470 |
747 |
16,7 |
P>0,10 |
|
Pe5 05 |
3,02 |
6 |
32 |
20 |
93 |
138,3 |
173,9 |
312,2 |
5604 |
88,5 |
1,6 |
P>0,10 |
|
Pe1 07 |
2,8 |
24 |
52 |
27 |
46 |
168,4 |
125,2 |
293,6 |
3717,6 |
302 |
8,1 |
0,000 |
|
Pe2 07 |
2,8 |
24 |
52 |
27 |
103,5 |
168,4 |
125,2 |
293,6 |
4511,2 |
755 |
16,7 |
0,004 |
|
Pe3 07 |
2,8 |
24 |
52 |
27 |
103,5 |
114 |
299 |
413 |
3909 |
208 |
5,3 |
0,004 |
|
Arr 07 |
3,03 |
17 |
60 |
30 |
123 |
142 |
285 |
427 |
5179 |
140 |
2,7 |
P>0,10 |
|
Ww 07 |
3,83 |
20 |
112 |
23 |
93 |
133 |
150 |
283 |
5499,5 |
637 |
11,6 |
P=0,05 |
|
Promedio |
|
|
|
|
|
|
|
|
|
283,2 kg |
6,4 % |
8 de 18 P<0,10 |
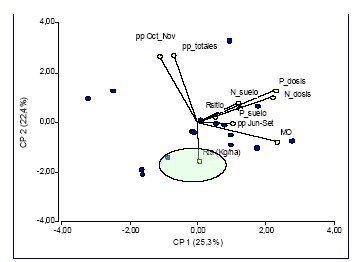
Figura 1: Respuesta absoluta a la inoculación con Pseudomonas fluorescens (Rta kg/ha) y su asociación negativa con las precipitaciones durante el período junio-noviembre y octubre-noviembre
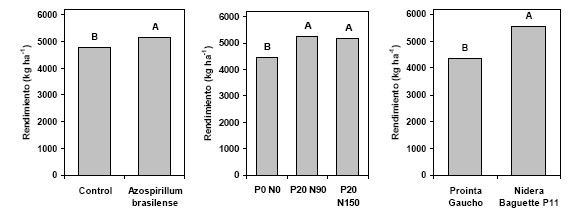
Figura 2: Producción media de trigo según tratamientos de semillas con Azospirillum brasilense, estrategias de fertilización NP y genotipos en Pergamino. En cada figura, letras distintas sobre las columnas muestran diferencias significativas (p<0,05). Tomado de Ferraris et al., 2008
Pseudomonas spp
Las Pseudomonas son otro amplio género bacteriano, en el cual se encuentran
especies con potencialidad para ser considerados PGPM. Han sido utilizadas con
fines agronómicos en nuestro país P. fluorescens y P. chlororaphis, en ese orden
de importancia. Los efectos atribuidos a este grupo bacteriano pueden resumirse
en una acción de biocontrol, la secreción de sustancias inductoras y la
solubilización de nutrientes.
La capacidad de ser agentes de biocontrol (Haas & Défago, 2005), se produce a
través de la secreción de antibióticos (i.e. pirrolnitrina, pioluteorina), la
inducción de resistencia sistémica en la planta y el agotamiento de elementos
esenciales para el crecimiento de hongos y bacterias patogénicas, producida por
la liberación al medio de pigmentos fluorescentes que actúan como agentes
quelantes, cuando estos elementos se tornan escasos en la rizósfera.
El control biológico no es sólo dependiente de las capacidades bacterianas, sino
también de la especie vegetal y la práctica de cultivo. En trigo, se ha
estudiado el efecto de decaimiento del pietín (“take-all decline”) por
monocultivo, descripto exhaustivamente por grupos del USDA (Cook, 2007). A lo
largo de décadas de estudio de las poblaciones de Pseudomonas asociadas a lotes
de trigo, se ha establecido una firme correlación entre la supresión del pietín
y la preponderancia de poblaciones de Pseudomonas productoras de los
antibióticos diacetilfloroglucinol y fenazinas (Cook, 2007). En nuestro país,
todavía no disponemos de datos sobre el efecto del cultivo y de las rotaciones
sobre las poblaciones de Pseudomonas y su posible correlación con la incidencia
de enfermedades.
Otro de los efectos favorables residiría en la producción de fitohormonas como
auxinas y giberelinas, a la vez que se reducirías niveles de etileno producido
ante situaciones de estrés moderado, especialmente estrés hídrico.
Por último a las Pseudomonas se les atribuye la capacidad de producir enzimas
fosfatasas, ácidos orgánicos (i.e. acido glucónico, cítrico) e inorgánicos (i.e.
ácido sulfhídrico, nítrico, carbónico) que por medio de la rotura de enlaces y
la acidificación del medio, incrementarían la recuperación del fósforo nativo
del suelo y la adquisición del aportado por fertilización.
Ensayos pioneros realizados con Pseudomonas en las localidades de Pergamino y
Chivilcoy durante los años 2000, 2001 y 2002, mostraron incrementos medios de
rendimiento de 310 kg ha-1 (García y Bach, 2003). En este grupo de seis ensayos,
los resultados más consistentes se observaron cuando la inoculación fue
acompañada de una adecuada fertilización con N y P. En el mismo sentido, en un
grupo de 11 experimento realizados durante 6 campañas agrícolas en el
centro-norte de Buenos Aires y Sur de Santa Fe, se cuantificó una respuesta
media a la práctica de inoculación con Pseudomonas de 294 kg ha-1, lo cual
representa un incremento de 8,4 % (Figura 3). Cada uno de estos sitios
representa a su vez, el promedio de diferentes estrategias de fertilización, con
o sin agregado de NP. En estos ensayos, como promedio, la eficiencia de uso de N
(EUN) pasó de 47 a 51 kg trigo/ kgN y la EUP de 181 a 195 kg trigo/ kgP, para
tratamientos testigo e inoculado, respectivamente.
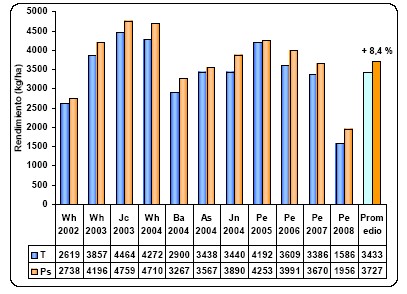
Figura 3: Rendimiento (kg ha-1) de tratamientos testigo e inoculados con Pseudomonas fluorescens en trigo entre los años 2002 y 2007 en el centro-norte de Buenos Aires y sur de Santa Fe. Las abreviaturas representan localidades Wh (Wheelwright, Santa Fe), Jc (Juncal, Santa Fe), Ba (Baigorrita, Buenos Aires), As (Ascensión, Buenos Aires), Jn (Junín, Buenos Aires), Pe (Pergamino, Buenos Aires) y el promedio de todas las localidades (Prom).
La asociación entre respuesta agronómica y variables de suelo y cultivo muestra condimentos interesantes (Figura 4) Así, en el eje 1 (horizontal) se observa que la respuesta relativa (%) se asocia de manera positiva a los rendimientos y la dosis de fertilizante fosforado, y negativamente con parámetros químicos de suelo como el nivel de P (Bray I), nitratos y MO. En el eje vertical se visualiza una relación directa entre respuesta y dosis de N aplicado, e inversa con las lluvias durante el ciclo. Considerando estos resultados, la respuesta a la inoculación con Pseudomonas spp sería más probable en ambientes empobrecidos, con bajo nivel de P y otros nutrientes, pero que comienzan a ser mejorados por buenas prácticas de cultivo y adecuada fertilización. Condiciones de estrés hídrico acentuarían estos efectos.
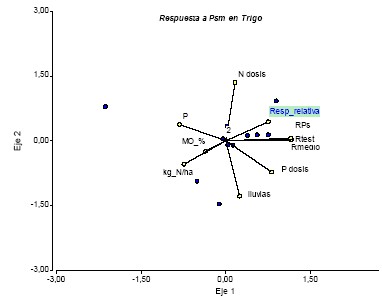
Figura 4: Respuesta relativa (%) a la inoculación con Pseudomonas fluorescens en trigo y su asociación positiva con los rendimientos del sitio (Rmedio), y negativa con las lluvias junio-noviembre (lluvias) y la fertilidad inicial del suelo (kg N/ha, P, MO%).
Micorrizas:
Las micorrizas son hongos ampliamente distribuídos en el suelo, capaces de
establecer asociaciones mutualistas con los vegetales. En la naturaleza, el 95 %
de las plantas las poseen (Trappe, 1977). Desde hace tiempo se conoce su efecto
favorable en especies perennes y forestales, pero últimamente se ha indagado su
utilidad en cultivos anuales.
Se reconoce de las Micorrizas su capacidad para mejorar la estructura del suelo
gracias al crecimiento del micelio y la secreción de glomalinas (Faggioli et
al., 2008). Asimismo, actuan como una prologación del sistema radicular
(Peterson et al, 2004)(Figura 5), facilitando la adquisición de agua y
nutrientes de baja movilidad como potasio (K), zinc (Zn) y especialmente P. Al
incrementar el flujo de P a la raíz, de manera indirecta se mejoran otros
procesos fisiológicos en que participa este nutriente. Por su menor diámetro,
las micorrizas tienen mayor superficie de absorción que las raíces del vegetal.
Si bien utilizan P bajo las mismas formas que las plantas, tienen mayor afinidad
por el nutriente y una concentración crítica en solución más baja para lograr su
absorción (García et al., 2006).
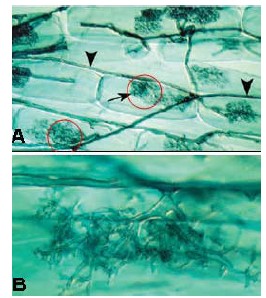
Figura 5: Proliferación de Micorrizas en células radiculares (Tomado de Peterson et al., 2004)
La proliferación e importancia agronómica de las micorrizas es más relevante
en suelos deficientes de P (Covasevich et al., 1995). Ante situaciones de
carencia, contribuyen con la síntesis de proteínas de estrés en planta. El
estrés conduce a la expresión diferencial de la información genética,
produciendo cambios en la síntesis de nuevas proteínas, llamadas micorrizinas,
las cuales posiblemente dotan a las plantas con la capacidad de adaptación al
estrés. La respuesta agronómica en rendimiento podría estar asociada a suelos
con baja disponibilidad de P, pero no se ha visto afectada por la dosis de
fertilizante agregado (Ferraris y Couretot, 2008).
Las Micorrizas necesitan oxígeno para vivir, por ello las poblaciones son muy
bajas en suelos de drenaje pobre y anegables. También en suelos salinos y/o
sódicos (Abbot y Robson, 1991). En cambio un suelo poroso y bien estructurado
las favorece. Cultivos de cobertura aumentan mucho la micorrización, lo mismo
que la siembra de maíz y sorgo, que tienen alta dependencia micorrítica e
incrementan su población (Faggioli et al., 2008). Las labranzas rompen el
entramado de micelios del hongo, destruyendo el efecto benéfico sobre la
estructura del suelo (Schalamuk et al., 2006). Dosis medias de fertilizante no
afectan a las micorrizas, al igual que insecticidas y herbicidas, a dosis
normales, (Coyne, 1999). Se ha mencionado que los funguicidas aplicados sobre
semilla pueden ser muy tóxicos para las micorrizas. En Argentina se han
difundido inoculantes comerciales con micorrizas en su formulación. Algunas
variantes tecnológicas de utilización están siendo estudiadas (Figura 6).
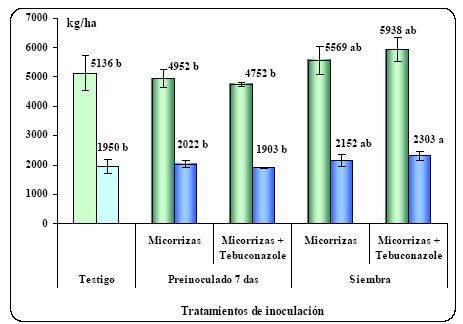
Figura 6: Materia seca a cosecha y rendimiento de grano de parcelas testigo o
inoculadas con micorrizas. Se evalúan momentos de inoculación y el uso conjunto
de fungicidas curasemillas. Pergamino, año 2008.
Bibliografía citada
Abbot L.K y A.D. Robson. 1991. Factors influencing the occurrence of
vesicular-arbuscular mycorrhizas. Agriculture Ecosystems and Environment.
35:121-150.
Caballero-Mellado, J., M.G. Carcano-Montiel, M.A. Mascarua-Esparza. 1992. Field
inoculation of wheat (Triticum aestivum) with Azospirillum brasilense under
temperate climate. Symbiosis, 13: 243-253.
Covasevic, F., H. Echeverría y Y. Andreoli. 1995. Micorriación vesículo-arbuscular
espontánea en trigo en función de la disponibilidad de fósforo. Ciencia del
Suelo 13:47-51.
Cook, R.J. 2007. Tell me again what it is that you do. Annual Reviews of
Phytopathology, 45:1-23.
Coyne M. 1999. Soil Microbiology: An exploratory approach. Delmar Publishers.
462 pag.
Díaz-Zorita, M., M.V. Fernádez-Canigia. 2008. Field performance of a liquid
formulationof Azospirillum brasilense on dryland wheat productivity, Eur. J.
Soil Biol. doi:10.1016/j.ejsobi.2008.07.001
Díaz-Zorita, M., R.M. Baliña, M.V. Fernández-Canigia, A. Perticari. 2004. Field
inoculation of wheat (Triticum aestivum L.) and corn (Zea mays L.) with
Azospirillum brasilense in the Pampas region, Argentina. RELAR, Rio de Janeiro
(Brasil).
Döbereiner, J., I. Marriel and M. Nery. 1976. Ecological distribution of
Spirillum lipoferum Beijerinck. Can J Microbiol 22: 1464-1473.
García, F.O.; L.I. Picone y A. Berardo. 2006. Fósforo. Pág. 99-121. En:
H.E.Echeverría y F.O. García (eds.) Fertilidad de Suelos y Fertilización de
cultivos.Editorial INTA, Buenos Aires, Argentina. 521p.
Faggioli, V., G. Freytes y C. Galarza. 2008. Las micorrizas en trigo y su
relación con la absorción de fósforo del suelo.
Ferraris, G. y L. Couretot. 2008. Evaluación de la inoculación con Micorrizas
bajo diferentes ambientes de fertilidad. pp 48-52. En: Trigo. Resultados de
Unidades Demostrativas. Proyecto Regional Agrícola.
Ferraris, G., L.. Couretot y M. Díaz Zorita. 2008. Respuesta de trigo a
tratamientos con Azospirillum sp. según niveles tecnológicos. CD Rom. VII
Congreso Nacional de Trigo.V Simposio Invernal de Cereales de siembra Otoño –Invernal.I
Encuentro del Mercosur.
García, R. y T. Bach. 2003. Efecto de la inoculación con Pseudomonas sobre el
rendimiento de trigo. Informe técnico 324, INTA - INSTITUTO NACIONAL DE
TECNOLOGÍA AGROPECUARIA. Centro Regional Buenos Aires Norte Estación
Experimental Agropecuaria Pergamino. 19 pp.
Haas D, Défago G. Biological control of soil-borne pathogens by fluorescent
pseudomonads. Nat Rev Microbiol. 2005 Apr;3(4):307-19. Review.
Okon Y. and C. Labandera-Gonzalez. 1994. Agronomic applications of Azospirillum:
An evaluation of 20 years worlwide field inoculation. Soil Biol. Biochem. Vol 26
(12):1591-1601.
Peterson R.L., H.B. Massicotte y L.H. Melville . 2004. Arbuscular mycorrhizas.
En: Mycorrhizas: Anatomy and Cell Biology. NRC-CNRC. Research Press.Otawa.
Canada. Chap.3: 57-79
Puente, M. y A. Perticari. 2006. Inoculación de trigo con Azospirillum. Trigo en
Siembra Directa. 97-99. Revista Técnica de la Asociación Argentina de
Productores en Siembra Directa, AAPRESID.
Russo A, Felici C, Toffanin A, Götz M, Collados C, Barea JM, Moënne-Loccoz Y,
Smalla K, Vanderleyden J, Nuti M (2005) Effect of Azospirillum inoculants on
arbuscular mycorrhiza establishment in wheat and maize plants. Biol Fertil Soils
41:301–309
TRAPPE, J. M. (1977). Selection of fungi for ectomycorrhizal inoculation in
nurseries. Ann. Rev. Phytopathol., 15: 203-222.
Schalamuk S, S. Velásquez, H. Chidichimo y M. Cabello. 2006. Fungal spore
diversity of arbuscular mycorrhizal fungi associated with spring wheat: effects
of tillage. Mycologia 1: 22–28.

